Primeras formas de vida
LUCA y el gran misterio del Origen
La escasez de información geológica y paleontológica durante las primeras fases de formación de la Tierra hace que la cuestión sobre el origen de la vida continúe siendo un misterio. Sin embargo, es una cuestión tan crucial que ha recibido grandes esfuerzos de investigación de todo tipo que intentan modelizar mediante datos indirectos, cómo serían tanto las condiciones de partida como las características de los primeros sistemas vivos. Podríamos concretar este “evento” como el de la aparición de la “primera célula” que representa la superación de un mundo prebiótico y la consecución de una independencia metabólica y funcional respecto de su entorno. Aquí intentaremos aclarar algo sobre ese momento de aparición y las características básicas de esta primera célula que ha sido denominada LUCA (Last Universal Common Ancestor).
Cuando sucedió
Desde hace décadas se discuten las posibles evidencias de vida en las rocas más antiguas. Es conocido el tópico de que en Akilia (Groenlandia) existen pruebas de vida de hace 3,83 Gy. Sin embargo, el fuerte metamorfismo que afecta a estas rocas hace que los datos sean difíciles de interpretar y que incluso a la luz de nuevos estudios cada vez más precisos, las conclusiones sigan siendo ambiguas y no concluyentes (Lapot, 2020).
Según los datos que tenemos hasta este momento, los registros claramente incontrovertibles de vida no aparecen realmente hasta el intervalo de los 3,5-3,4 Gy y provienen principalmente del Cratón Arcaico de Pilbara en Australia. Así, los primeros fósiles químicos claramente biogénicos los encontramos en la formación Dresser con 3,47 Gy y consisten en firmas isotópicas de azufre biogénico. Por otro lado, los primeros microfósiles y estromatolitos verdaderos provienen de la Formación Strelley Pool, con 3,4 Gy.
Estas evidencias fósiles hablan de una biota ya diversificada. Por tanto, LUCA habría surgido con anterioridad. Y aquí ya entramos en el campo de la especulación.
La evolución prebiótica necesaria para la aparición de LUCA necesita de un entorno relativamente estable. En este sentido el “Gran Bombardeo Tardío” o GTB que habría tenido lugar a finales del Hadeense (4100-3900 m.a.), es un inconveniente ya que aniquilaría cualquier avance de una posible biosfera.
Si tenemos en cuenta este evento, la aparición de LUCA tendría que situarse en un intervalo de tiempo posterior, desde los estertores del GBT (3900 ma) hasta la primera evidencia fósil claramente establecida (los 3,4 Gy de Pilbara).
Sin embargo, en algunos trabajos se propone que el GBT no habría sido lo suficientemente intenso como para detener o borrar la evolución prebiótica y que ésta habría aparecido mucho antes, en algún momento posterior al de la formación del sistema Tierra-Luna (Abramov y Mojzsis 2009). En este mismo sentido, otros autores proponen incluso que el GTB no habría llegado a suceder y que es fruto de una interpretación muy simplista de los datos obtenidos de las misiones lunares (Boehnke & Harrison, 2016).
Como vemos, en ambos casos, la aparición de LUCA sería muy antigua, en algún momento entre los 4200 y 3900 m.a. Hallazgos más recientes de posibles microfósiles con una edad entre 4280 y 3760 m.a. encontrados en Nuvvuagittuq, Canadá (Dodd et Al. 2017), indican también un origen en el Hadeense. Sin embargo, será muy difícil demostrar su biogenicidad (Lapot, 2020).
Como se ve, la fecha para el surgimiento de LUCA presenta dos posibles soluciones. Por un lado, para la hipótesis Hadeense, cogeríamos algún punto intermedio entre 3900 y 4200 m.a. relativizando la importancia del GTB. Por otro lado, en la hipótesis Arcaica, el punto intermedio se escogería entre los 3900 y los 3500 m.a.
Esta discusión, aunque de momento deja una gran ambigüedad, es muy fructífera ya que promueve una increíble mejora de los métodos analíticos y el aumento de la precisión en las dataciones que se ha ido plasmando en un avance acelerado de la investigación.
IDENTIDAD DE LUCA
La comparación de las principales ramas de organismos, es decir, bacterias, arqueas y eucariotas, nos muestra que todos tendrían un origen común. Esta es la idea básica que subyace al concepto de LUCA y que nos acompaña desde los tiempos de Darwin.
Hoy en día, la Proteómica comparada permite encontrar los genes y proteínas ancestrales y nos permite perfilar cómo sería este organismo (Gabaldón 2009; Lane y Martín, 2012; Sojo et al. 2014):
1. Sabemos que tendría toda la maquinaria de transcripción y traducción génica con un código genético ya fijado.
2. Metabólicamente poseería gluconeogénesis y parte del ciclo de Krebs como base para las rutas de síntesis molecular y una fijación de CO2 mediada por proteínas férricas denominadas ferredoxinas. Esto significa que sería anaerobio y quimioautótrofo. es decir, que viviría en un medio sin oxígeno y que obtendría la energía de compuestos inorgánicos reducidos (por ejemplo, el H2) en lugar de obtenerla de la luz.
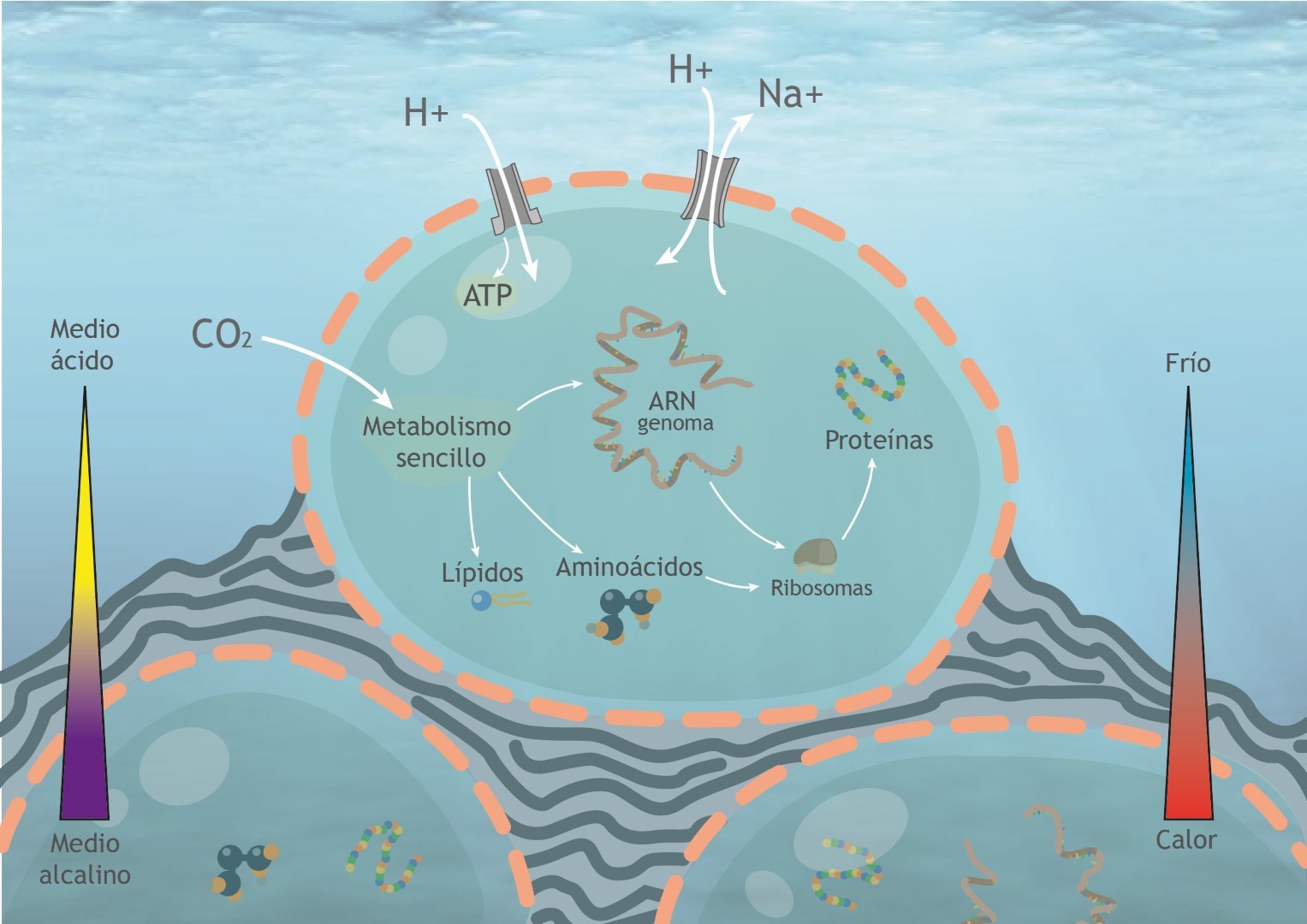
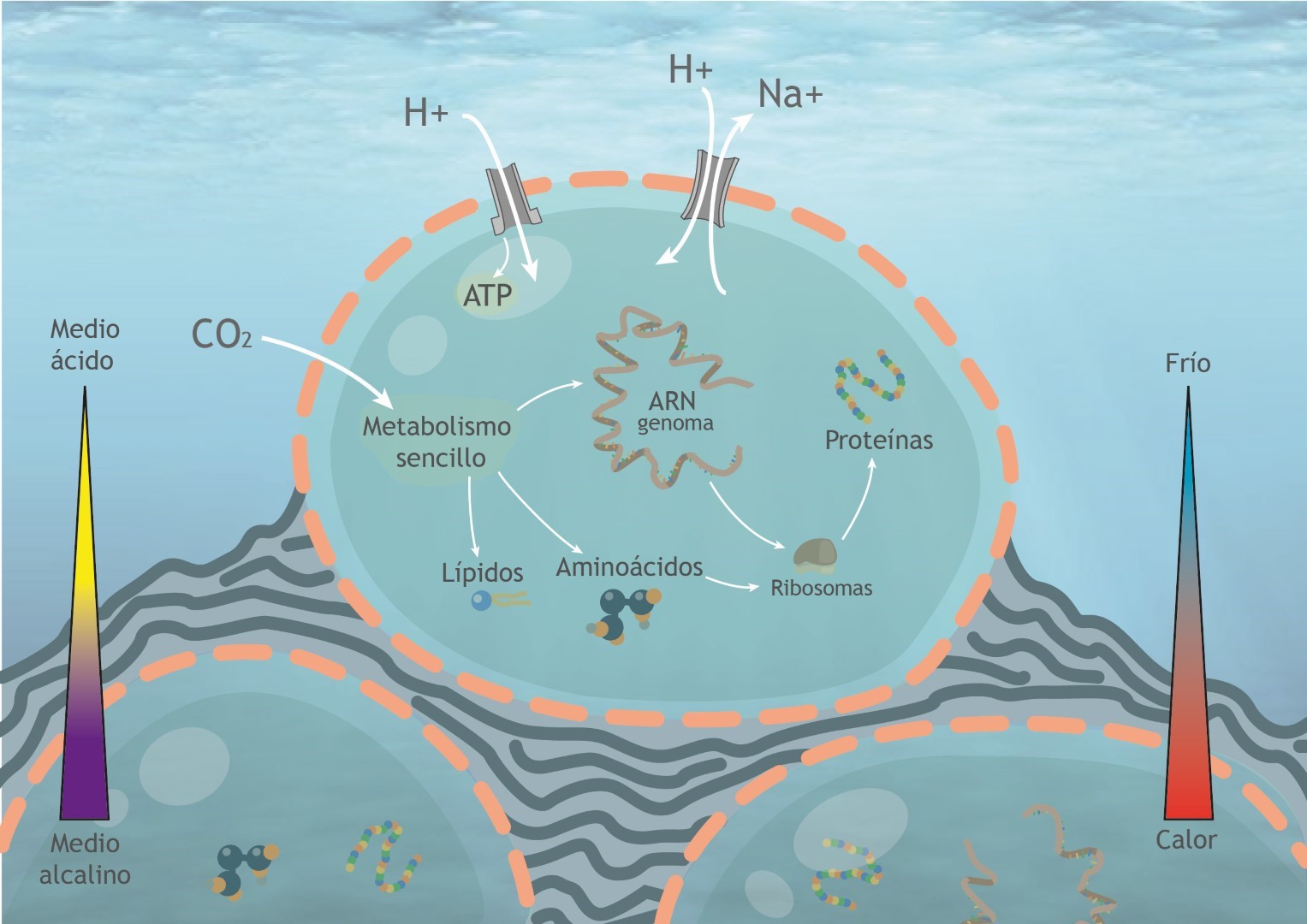
3. La membrana, aunque ya genéticamente codificada, sería altamente porosa, hecha de ácidos grasos simples y probablemente asociada a finas películas inorgánicas de hierro-azufre. Esta membrana aún no tendría capacidad quimiosmótica aunque ya poseería proteínas de membrana de dos tipos, ATPasas que aprovechan el flujo de protones a su través para sintetizar ATP, y proteínas “antiporte” de intercambio, que transportan iones sodio hacia fuera por protones hacia el interior, lo que mejora la eficiencia del flujo de protones. Este tipo de membranas harían a LUCA aún dependiente de los gradientes químicos externos y por tanto solamente viable en entornos hidrotermales adecuados donde existen estos gradientes.
4. Por último: según parece, en LUCA no estarían presentes los elementos necesarios para la replicación del ADN y se especula si su genoma aún sería de ARN, utilizando ADN a modo de paso intermedio, mediante enzimas como la transcriptasa inversa típica de los retrovirus. La célula se dividiría simplemente por crecimiento y rotura. Sólo más tarde, en el camino hacia la independencia quimiosmótica, aparecería un nucleoide de ADN más estable.
Fósiles químicos
Los fósiles químicos son concentraciones de isótopos, iones o moléculas que sólo pueden ser explicados mediante un proceso biogénico llevado a cabo por organismos vivos. En la cuestión del origen de la vida, el fósil químico más importante se refiere a la presencia de una relación característica entre dos de los isótopos estables del carbono, el C12 y el C13. Todos los seres vivos incorporan en sus moléculas orgánicas prioritariamente C12. Por tanto, se puede distinguir cuándo la materia carbonosa tiene origen biogénico estudiando si la relación de C12/C13 es la adecuada.
Quimiosmósis
Una de las características esenciales de las células y por tanto del fenómeno de la vida se basa en la propiedad de la quimiosmósis. Mediante la quimiosmósis la célula es capaz de bombear de forma activa iones y moléculas de dentro hacia afuera de las membranas, creando con ello gradientes que utiliza para producir energía en forma de ATP, cuando estos compuestos regresan a la célula por canales especiales con esta función (ATPasas). Mediante este bombeo activo, las células posteriores a LUCA (arqueas y bacterias) fueron capaces de independizarse del medio hidrotermal y conquistar todos los ambientes de la superficie terrestre.
Qué sucedió
Este modelo de LUCA basado en la Proteómica comparada sitúa a nuestro ancestro anclado aún a un ambiente hidrotermal. Pero ¿cómo llegó a aparecer? Se han ido encontrando de modo experimental algunos fragmentos del proceso que plantean los siguientes pasos:
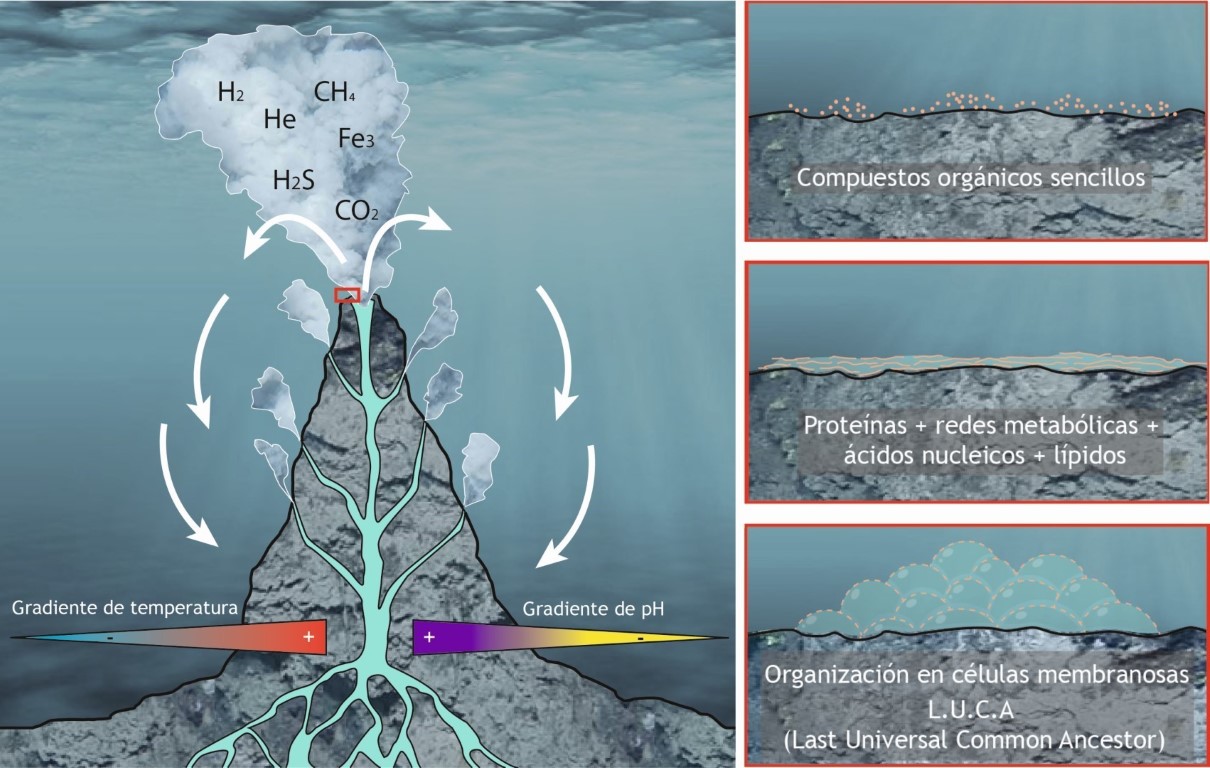
1. SOPA INORGÁNICA PRIMORDIAL. Las moléculas pre-orgánicas simples se forman de modo espontáneo en varios entornos donde existen gradientes físico-químicos adecuados. Estos entornos se han encontrado incluso en meteoritos y cuerpos cometarios y hay muchas posibilidades propuestas, entre las que destacan las de naturaleza hidrotermal. Estas moléculas serían amoníaco, hidrógeno, nitrito, sulfuro, el sistema hierro-azufre, formas de fosfato solubles, metano, metanol, acetileno, formaldehído, formiato, etanol, acetato, gliceraldehído, cianuro y aminoácidos sencillos.
2. CATÁLISIS INORGÁNICA. Además, parece que es también clave que este sistema de moléculas esté suficientemente concentrado y en presencia de situaciones que faciliten las reacciones. Esta facilitación catalítica provendría en principio de las superficies minerales (sulfuros, minerales de la arcilla y otras varias propuestas), así como de la existencia de iones metálicos (Fe, Cu, Mn, Ni, Zn, Mo, etc…).
3. PEQUEÑOS PÉPTIDOS. Según los modelos más modernos y tras muchos años de desarrollo del “mundo de ARN” hoy día algo agotado, parece que se va reconociendo que el primer salto importante está en el desarrollo de los primeros péptidos a partir de cadenas pequeñas de aminoácidos inorgánicos. Esta es la hipótesis que podemos llamar de “proteínas primero” (Dill y Agozzino, 2021) y que es una formulación más moderna del grupo de teorías denominadas “Metabolismo primero”. Estos péptidos parecen formarse de forma espontánea en determinadas condiciones.
4. PLEGAMIENTO PEPTÍDICO Y DESARROLLO DE FUNCIÓN. Los péptidos, a medida que se van haciendo más largos y debido al carácter hidrófobo de algunos de ellos, comienzan a plegarse. El plegamiento confiere morfología y capacidad catalítica. La catálisis tiene dos aspectos: uno la autocatálisis, que promueve la multiplicación del péptido y otro la catálisis de una reacción sobre algún sustrato determinado del entorno que promueve que determinadas reacciones comiencen a aumentar su velocidad. Además, estas proto-proteínas se asociarían fácilmente a los catalizadores inorgánicos (iones metálicos).
5. ACOPLAMIENTO DE REACCIONES. Algunas reacciones acaban acoplándose ya que el producto de una es el sustrato de la otra. Esto promueve la aparición de cadenas de reacciones (proto-metabolismo) y por otro lado el aumento en la complejidad y longitud de los péptidos.
6. MEMORIA: ÁCIDOS NUCLEICOS. En estos estadios del desarrollo y en paralelo comenzarían a sintetizarse los nucleótidos. En los experimentos de laboratorio es necesaria la intervención de la radiación UVA y esto es un problema ya que implica necesariamente un ambiente expuesto a la radiación solar y lejos del ambiente hidrotermal submarino. Una solución a este problema es un ambiente hidrotermal desarrollado en la superficie, una especie de laguna hidrotermal (Mulkidjanian et al. 2012).
7. COMPARTIMENTACIÓN. Estas redes de reacciones pueden estar concentradas en algún tipo de compartimento inorgánico del ambiente hidrotermal, ya sean películas inorgánicas de hierro-azufre y/o pequeñas vesículas lipídicas porosas (Coacervados).
8. CÓDIGO GENÉTICO. La asociación entre proteínas y nucleótidos proviene de la afinidad natural entre las bases nitrogenadas y los aminoácidos ya que son compuestos que se caracterizan por su capacidad para enlazarse por puentes de hidrógeno. Esta asociación electroquímica promovería que la información de la secuencia de los péptidos quedara encriptada en moléculas de nucleótidos complementarios, dando lugar al código genético. Poco a poco comenzaría a formarse una capacidad de repetición de las redes metabólicas a través de la encriptación en cadenas de nucleótidos de la información de sus proteínas catalizadoras y del orden de su formación. Pero los nucleótidos también tienen capacidad catalizadora (ribozimas), ¿por qué no fueron ellos los que desarrollaron la creación de las redes metabólicas? La respuesta puede ser esta: la capacidad de combinación de los 20 aminoácidos produciendo largas cadenas reactivas con capacidad de plegamiento hacen posible un elenco casi infinito de formas funcionales, asociadas a catalizadores metálicos inorgánicos, muy superior y más estable químicamente que el que se consigue con cadenas más o menos largas y plegadas de nucleótidos de ARN.
9. EVOLUCIÓN. La memoria promovida por la encriptación en ARN permite que comience a actuar la evolución (selección natural, simbiosis, fusión, etc.) de redes de reacciones energéticamente acopladas que son capaces de copiarse a sí mismas y que quedan protegidas por una membrana. Todo ello acaba por dar lugar a un organismo equivalente a LUCA tal y cómo lo hemos descrito más arriba.
Dónde sucedió
Respecto al entorno en el que debió producirse el desarrollo prebiótico y como ya se ha sugerido, las propuestas más recientes se decantan por los respiraderos hidrotermales. En principio los entornos hidrotermales más lógicos serían oceánicos. Primeramente, se pensó en los humeros negros, pero los gradientes de energía de las fumarolas volcánicas submarinas son excesivos, el pH es ácido y la estabilidad en el tiempo escasa.
Un modelo más adecuado lo proporciona el modelo de la “Ciudad Perdida” (Bradley, 2010). Está basado en los humeros blancos que se producen en los procesos de serpentinización de la corteza oceánica. Este proceso es la transformación de olivino en otro mineral denominado Serpentina y ocurre de forma natural bajo condiciones hidrotermales, dando, como producto final, además de la serpentina, hidrógeno molecular (H2). Estos humeros blancos se sitúan algo más distales de la zona volcánica y por tanto en un entorno de temperaturas más bajas y que además son alcalinos y mucho más estables en el tiempo.
La aparición y desarrollo de los ácidos nucleicos hace que necesitemos además radiación UVA sólo presente en ambientes superficiales. Este desacoplamiento entre el modelo hidrotermal submarino y los nucleótidos sólo ha sido respondido con la propuesta de un ambiente hidrotermal superficial (Mulkidjanian et al., 2012).
La conexión entre ambos sub-ambientes hidrotermales es actualmente una cuestión no resuelta.
Cómo lo sabemos
La investigación sobre el origen de la vida es muy compleja y abarca muchas disciplinas. El marco general lo ofrece la geología y la petrología, pero los datos específicos provienen de la paleontología, geoquímica, bioquímica, astrobiología, etc…
Son especialmente importantes los datos referidos a las dataciones de las rocas (datos isotópicos), los datos para interpretar la biogenicidad de los fósiles supuestos, que suelen ser bioquímicos, de microscopía avanzada y espectroscópicos (espectroscopía Raman) además del estudio de las relaciones de isótopos estables. La capacidad de aplicar estos análisis en zonas microscópicas de las muestras es el mayor reto, que en la actualidad es ya resolutivo en algunos casos hasta la nanoescala.
Por otro lado, son esenciales los experimentos en el laboratorio de síntesis de biomoléculas y rutas metabólicas, así como estudios evolutivos relacionados con el genoma y el proteoma en la búsqueda de proteínas y genes que sean ancestrales.
Sabías que ...
Nos preguntamos: ¿Existe un lugar para los virus en el árbol de la vida? Los virus no pueden haber existido antes que las células ya que necesitan sus mecanismos de replicación, es decir, necesitan un huésped. Pero existe una teoría (Forterre, 2005) que propone que los tres linajes celulares del árbol de la vida, todos ellos con ADN (Eubacteria, Arqueobacteria y Eukarya), se formaron con posterioridad a LUCA gracias a la aparición de virus de ARN a expensas de LUCA. Estos virus tienen una velocidad de mutación muy alta y se piensa que a partir de ellos surgirían enzimas capaces de sintetizar desoxirribosa, que es el azúcar que diferencia el ARN del ADN. Por tanto, de los virus de ARN habrían aparecido los virus de ADN y posteriormente de ellos, el ADN procariota. Todo ello podría explicar la sorprendente coexistencia de dos maquinarias de replicación de ADN: una para Bacterias, la otra para Eukarya/Archaea.
