El origen de la célula eucariota
Quimeras e individualidad
La vida procariota es quizás la esencia de la vida en la Tierra ya que está presente desde los comienzos y aparece en todos los entornos conocidos de la biosfera. Por eso, cabe preguntarse por qué aparecieron los organismos Eucariotas, formados por nuevas estructuras celulares gigantescas y mucho más complejas. Una de las respuestas más populares dice que estas nuevas células, al ser más grandes, podrían alimentarse sin problemas de las procariotas más pequeñas y obtener así una ventaja. Esto nos lleva a una pregunta aún más difícil: Si esto es así, ¿por qué tardaron tanto en triunfar desde el momento de su aparición hasta hacerse ubicuas y dominantes? ¿por qué no hay un registro de un Big-Bang eucariota? Los nuevos datos que vamos conociendo revelan que la historia debió de ser mucho más compleja y nos enfrenta a más preguntas que respuestas.
Cuando sucedió
Fósiles químicos provenientes de hidrocarburos de 2,7 Gy en Pilbara, Australia, han sido citados durante mucho tiempo (Brocks et al. 1999) como la primera evidencia de vida eucariota debido al tipo de lípidos identificados. Sin embargo, hoy en día se rechaza esta hipótesis ya que se ha comprobado que los hidrocarburos provienen de un proceso de contaminación con materiales más modernos (French et al. 2015).
Los primeros fósiles claramente identificados de organismos eucariotas tienen una edad aproximada de 1,65 Gy y provienen del norte de China (Miao et al., 2019) y Australia (Javaux, 2019). Consisten en pequeños microfósiles palinomorfos del grupo de los Acritarcos. Se identifican muy bien como eucariotas gracias a su tamaño y a la estructura compleja de sus paredes que hace pensar en la posesión de un citoesqueleto desarrollado y un sistema de endomembranas (y por tanto núcleo verdadero).
Existen hallazgos más antiguos, pero todos son dudosos. Entre ellos destacamos los fósiles de la formación Francesville en Gabón, por su antigüedad (2,1 Gy) y su excelente conservación. Son icnofósiles que consisten en moldes tubulares y laminares piritizados que han sido interpretados como fruto de la actividad de colonias de organismos ameboides quizás similares a hongos mucilaginosos, aunque de momento, de naturaleza eucariota no comprobable (El Albani et al, 2019).
Todos estos datos implican que el intervalo de aparición de los primeros Eucariotas abarca desde los 2,2 Gy a los 1,7 Gy., siendo imposible de momento una mayor precisión.
Acritarcos
Los acritarcos son microfósiles marinos obtenidos mediante técnicas de extracción de polen. Muchos de estos microfósiles son considerados como “microproblemática” aunque constituyen el único registro fósil regular durante el paleoproterozoico y Mesoproterozoico. Siguen siendo abundantes incluso durante el Paleozoico pero disminuyen drásticamente a partir de ahí. Muchos taxones pueden relacionarse con algas o bien con hongos y siempre dentro de los eucariotas unicelulares.
Qué sucedió
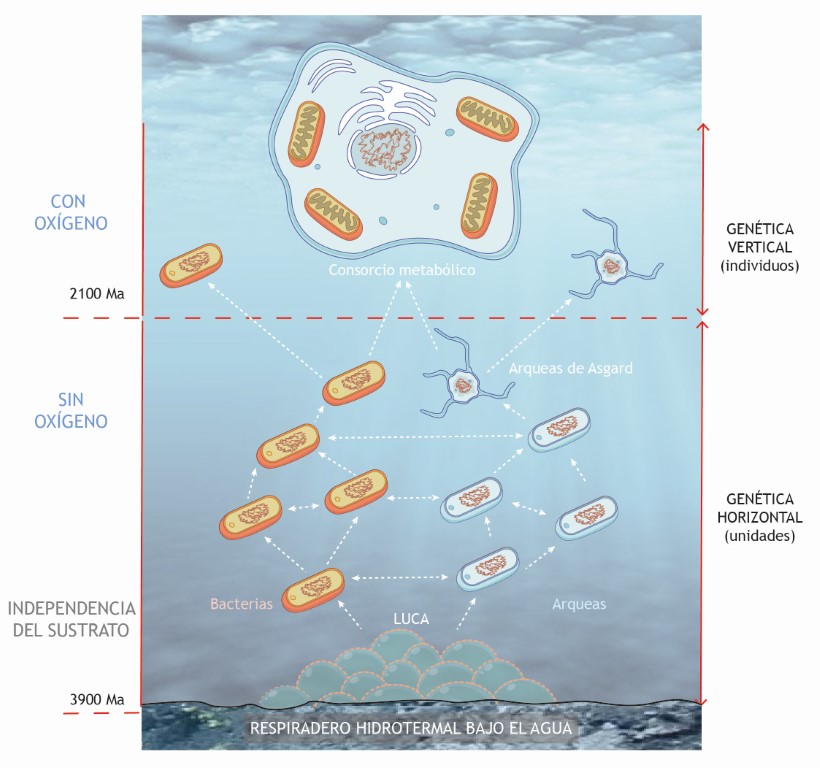
Para algunos autores, el evento GOE (la gran oxidación), ocurrido entre 2400 y 2100 millones de años, podría explicar este proceso como una respuesta adaptativa a una creciente concentración de oxígeno ambiental.
Los organismos de partida para una evolución hacia la célula eucariota podrían pertenecer a un grupo especial de arqueobacterias, las arqueas de Locki, dentro del grupo mayor de las arqueas de Asgard. Estas arqueas tienen una genética que es precursora de proteínas eucariotas como la actina, esenciales en el citoesqueleto y la construcción de endomembranas, además de otros muchos parecidos (Spang et al., 2015).
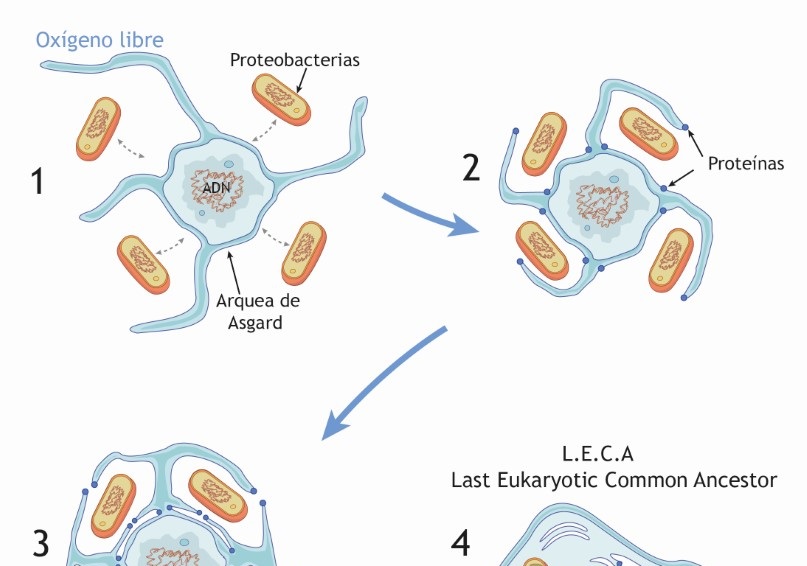
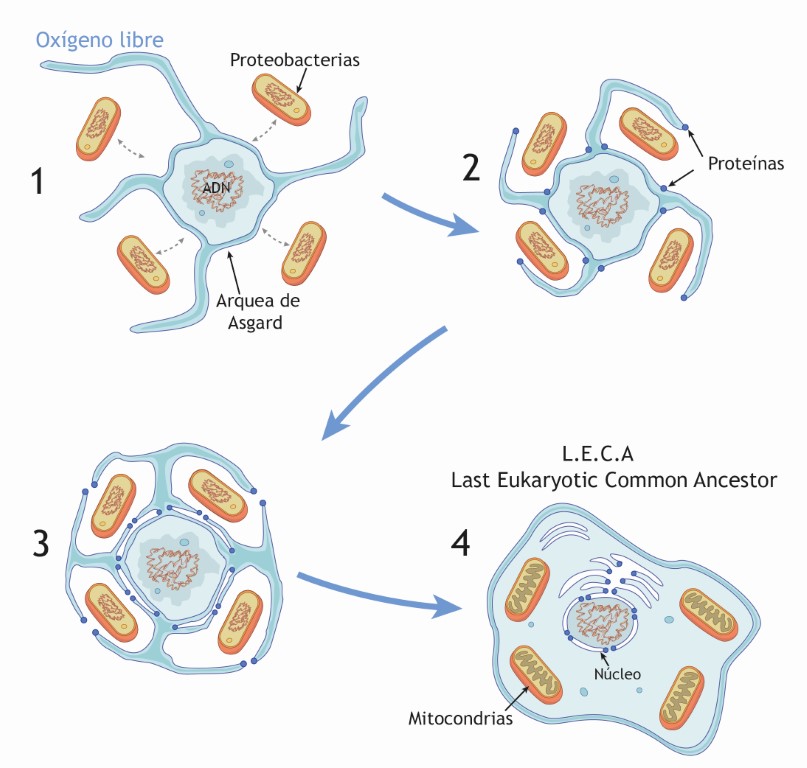
Recientemente se han cultivado cepas de este tipo de arqueas y se han investigado en su entorno natural. Su modo de nutrición es sintrófico, lo que significa que pertenecen a un consorcio de microorganismos con metabolismos complementarios que se apoyan mutuamente. En este contexto emerge un modelo que se ha denominado “entrelazar-engullir-endogenizar” que se resume en la figura (Imachi et al., 2020).
- Inicio. Al comienzo tenemos una asociación formada por un arquea anaeróbica (en azul y gris) que desarrolla cierta tolerancia al oxígeno al acoplar su metabolismo al de una bacteria sulforeductora (en naranja).
- Entrelazar. En este medio ambiente algo más oxigénico, que ha sido denominado ambiente de transición redox (López-García y Moreira 2022), puede suceder una ampliación del consorcio mediante la asociación no obligada con una alfa-proteobacteria primitiva (la futura mitocondria) lo que promueve una todavía mayor tolerancia al oxígeno.
- Engullir. Las capacidades de las expansiones membranales y el citoplasma del huésped arqueal “engullen” a la proto-mitocondria que queda englobada mientras que el nucleosoma arqueal es protegido por endomebranas.
- Endogenizar. A partir de aquí comienza un proceso ordenado de acoplamiento y evolución de los genomas de huésped y endosimbionte. Este proceso es el motor que hace avanzar el desarrollo de las endomembranas y del núcleo protegido. Finalmente surge LECA (Last Eukaryotic Common Ancestor), o sea la célula eucariota de la que descienden todas las células eucariotas actuales.
Este proceso se concibe como una adaptación a un entorno progresivamente más oxigenado y por tanto como respuesta al periodo GOE. En concreto, en medios de transición redox en zonas oceánicas poco profundas (¿quizás en esteras microbianas?), ya que se sabe que los fondos oceánicos más profundos fueron totalmente anóxicos (López-García y Moreira 2022).
Sin embargo, también es conocido, que dicho periodo dura aproximadamente unos 200 m.a. hasta el evento Lomagundi y después los niveles de oxígeno vuelven a bajar por causas aun escasamente investigadas quizás relacionadas con la tectónica global de aquella época (Lyons et al., 2014).
Esta bajada en los niveles de O2 podría explicar la ausencia de un big-bang eucariota, aunque los intervalos de tiempo implicados y los escasos y enigmáticos hallazgos fósiles no permiten sincronizar bien ni entender con una mínima precisión la secuencia de los sucesos.
Esta vuelta atrás en el contenido de O2 permanecería hasta finales del Proterozoico durante muchos cientos de millones de años. En este mundo escasamente cambiante de océanos predominantemente anóxicos, las innovaciones en el mundo eucariota fueron apareciendo lentamente hasta que, con las glaciaciones neo-proterozoicas, el oxígeno vuelve a dispararse, provocando una serie de eventos que aceleran la evolución, promoviendo, esta vez sí, la explosión del mundo de los protozoos, algas e incluso de los primeros animales.
Por qué sucedió
El salto cuántico entre células procariotas y eucariotas es enorme. La ausencia de grupos de organismos intermedios en cuanto a estructura entre ambos niveles de organización, ha dificultado mucho la investigación y han promovido una extensísima bibliografía plagada de todo tipo de teorías.
Una de las principales diferencias que aporta la aparición de un núcleo se refiere al modo en el que fluye la información genética entre las poblaciones. Este modo depende casi siempre de la reproducción y por eso decimos que es de transmisión vertical. La identidad genética de cada individuo está protegida y aislada. Sin embargo, en el mundo procariota el intercambio genético “horizontal” predomina, de modo que la identidad individual es algo mucho más relativo.
Se dice que las células eucariotas son quimeras. La integración de estas quimeras surge de las capacidades de intercambio genético horizontal en el mundo procariota. Pero el resultado final es todo lo contrario, el refuerzo de la individualidad protegida por un núcleo.
Resumiendo, la aparición de las “quimeras” eucariotas provendría de un mecanismo de refuerzo de consorcios microbianos y como respuesta metabólica a la oxigenación ambiental. Cuando los niveles de oxígeno volvieron a caer, las estirpes pudieron mantenerse en algunos nichos residuales. Su evolución fue restringida y lenta respecto a los entornos procariotas gracias a la recién descubierta individualidad y a su modo de propagarse a través de la reproducción. Este puede ser el porqué de que el salto cuántico entre eucariotas y procariotas sea tan grande: una evolución diferente durante los cientos de millones de años que duró el Proterozoico.
Cómo lo sabemos
La información utilizada para reconstruir la historia de la aparición del LECA se basa en gran medida en estudios de filogenómica, que es una especie de paleontología de genes y moléculas. A través de la identificación de los genes y las proteínas más primitivos y sus relaciones filogenéticas se ha logrado grandes avances, como es el de identificar qué grupo de arqueas son el grupo más cercano al primer huésped: las arqueas de Asgard.
A partir de este punto, el complejo estudio de la genómica y la ecología de estos organismos está dando las claves para proponer hipótesis cada vez más precisas que van incorporando parte de las hipótesis anteriores y desechando algunas de ellas.
Pero todavía estamos muy lejos de lograr integrar la filogenómica con los datos geológicos debido a la escasez de información y a la ambigüedad de muchos fósiles, en un periodo tan vasto como es el Proterozoico inferior y medio.
